Spletna revija za znanstvenike, strokovnjake
in nevroznanstvene navdušence
Naslovnica Članki Intervjuji Mnenja Zdravje Korenine eSinapsa Številke ![]()
Branje in možgani
letnik 2019, številka 16
uvodnik
članki
Maša Čater
Saša Koprivec
Infekcije osrednjega živčnega sistema s flavivirusi
Maja Potokar
Raziskava: Kako depresija vpliva na kognitivne sposobnosti?
Vida Ana Politakis
Razvoj depresije pri otrocih z vidika navezovalnega vedenja
Neža Grgurevič
Sonja Prpar Mihevc
Umetno inteligentna nevroznanost: srečanje nevronskih mrež in možganske fiziologije
Kristijan Armeni
Čebelji strup pri preventivi nevrodegenerativnih bolezni in priložnost za klinično prakso
Matjaž Deželak
intervju
Maša Čater
aktualno
Tina Giber
Teden možganov 2019: Klinika bolečina
Danaja Metul
kolofon
letnik 2019, številka 16
Otroci se v prvem razredu osnove šole učijo pisanja in prepoznavanja črk, nadaljujejo pa s povezovanjem le-teh v besede in nato več besed v stavke. Spoznavanje s črkami in zanimanje zanje pa se po navadi prične že mnogo prej. Prav vznemirljivo je, kako hitro otroci naredijo preskok v znanju. Samo nekaj tednov vaje je potrebnih in že se iz popolnega nebralca prelevijo v bralca. S časom se veščina branja še utrdi in izboljša. Ob učenju branja pa poteka kompleksen razvoj možganov.
Razvoj pismenosti
Branje in pisanje med drugim vključujeta mentalno predstavo nabora vidnih simbolov, ki so povezani v besede preko različnih fonoloških pravil. Potrebno je tekoče povezovanje med semantičnim besediščem, ki predstavlja shrambo pomena besed, fonološkimi predstavami, ki velja za shrambo slušnega zvoka besede, ter ortografskim besediščem, ki združuje znanje vidne podobe napisanih besed. Ti trije tipi besedišča niso podedovani, temveč se razvijejo kot posledica izpostavljenosti materinemu jeziku. Tako možgani niso genetsko pogojeni s samim razvojem pismenosti, ampak se z izpostavljenostjo vidnim in slušnim jezikovnim dražljajem učijo brati in pisati, kar pa zahteva reorganizacijo možganov 1.
Spremembe v možganih kot podlaga za razvoj bralnih sposobnosti
Branje velja za priučeno veščino, ki je močno odvisna predvsem od dveh parametrov; od razvitosti možganov ter bralnih izkušenj. Razvoj možganov poteka na podlagi sprememb v sivini in belini možganov. Bela možganovina sestavlja aksonske povezave v notranjosti možganov, medtem ko skorjo in možganska jedra v notranjosti poimenujemo kot sivino. Spremembe ob razvoju možganov so tako regionalno kot časovno specifične; v splošnem velja, da se volumen sivine zmanjšuje, medtem ko se volumen beline povečuje med prehodom iz otroštva v odraslost. V filogenetsko starejših regijah možganov pride do sprememb v volumnu možganovine pri razvoju možganov prej kot v filogenetsko novejših regijah. Spremembe v sivini in belini so pod vplivom tako genetike kot okolja. Genetika vpliva na razvoj delov možganov v različni meri. Pri otrocih in mladostnikih so ugotovili, da genetski faktorji v največji meri (80 %) vplivajo na celotni volumen možganov in v podobni meri (77% in 89 %) tudi posamično na volumen sivine in beline. Razlike v vplivu so celo med hemisferama, saj je volumen desne hemisfere bolj podvržen vplivu genov (v deležu 66 %) kot volumen leve (56 %) 2. Po drugi strani tudi izkušnje iz okolja močno vplivajo na razvoj možganov pri otrocih. Psihosocialni dejavniki iz okolja vplivajo na razvoj nevronov in na spremembe v določenih delih možganov. Potrjen je na primer vpliv materine podpore in bližine v predšolski dobi na hitrejšo rast hipokampusa pri otrocih v šolski dobi in kasneje v odraslosti. Bolj razvit hipokampus pa se nato odraža v bolje razvitem čustvenem razvoju, spominu in sposobnosti uravnavanja stresa 3. Z ozirom na razvoj sposobnosti branja lahko iz najnovejših dognanj povzamemo, da pri otrocih, ki so boljši bralci in ki berejo hitreje ter pogosteje kot nekoliko slabši bralci, prihaja do drugačnega razvoja določenih regij v možganih 4.
Z branjem povezani deli možganov
Z metodami slikanja možganov so dokazali, da branje omogoča in izvaja področje leve hemisfere, kjer pride do mreženja nevronov iz frontalnih, temporo-parietalnih in okcipito-temporalnih predelov skorje (slika 1). Ti omogočajo povezavo med vidnimi (ortografskimi) in slušnimi (fonološkimi) informacijami ter konceptualnimi (semantičnimi) podobami. Otroški možgani hitro vzpostavljajo nove sinaptične povezave, kar omogoča nove miselne sposobnosti. Pri tem prihaja do večje in natančnejše funkcionalne in strukturne povezanosti regij možganov med seboj. Slušni korteks, odgovoren za sprejem glasovnih predstav, se povezuje s predeli za procesiranje glasov – levi inferiorni frontalni girus 1.
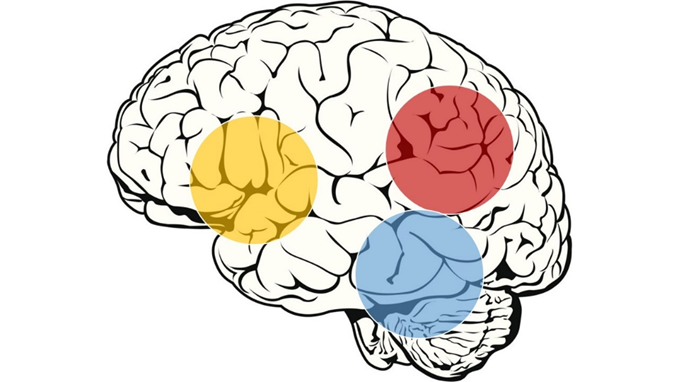 Slika 1: Anatomski deli možganske skorje leve hemisfere, ki so povezani z branjem. Rumena – inferio-frontalni del, rdeča – temporo-parietalni del, modra – okcipito-temporalni del.
Slika 1: Anatomski deli možganske skorje leve hemisfere, ki so povezani z branjem. Rumena – inferio-frontalni del, rdeča – temporo-parietalni del, modra – okcipito-temporalni del.
Disleksija in aleksija
Ena izmed motenj v učenju branja je nevrobiološka motnja disleksija. Tako pri otrocih kot odraslih so z nevrološkim slikanjem dokazali zmanjšano aktivnost levega temporo-parietalnega in okcipito-temporalnega predela skorje med fonološkim procesiranjem in branjem. Disleksični bralci pravzaprav izkazujejo povečano aktivnost desne hemisfere in bilateralnih prefrontalnih dorzalnih predelov, kar naj bi predstavljalo kompenzacijo za slabše delovanje levih posteriornih predelov 1. Disleksija velja za dedno nevrološko razvojno motnjo, ki se izraža kot konstantno, doživljenjsko oteženo branje in/ali črkovanje, ki pa ni povezana z nižjo inteligenco ali okoljskimi vplivi 5. Motnje branja se lahko kažejo na različne načine. Dislektiki pogosto težko prepoznavajo besede, ki jih sicer že poznajo, in jih težko črkujejo. Težave imajo pri pisanju, hitrem branju, izgovarjavi besed ter pri razumevanju pisane besede. Pogosto imajo disleksiki težave tudi s številkami. Dednost igra pomembno vlogo pri pojavnosti disleksije. Le-ta se pojavi pri otrocih, kjer je eden od staršev disleksičen, kar v 40-60 %. V kolikor sta disleksična oba starša, pa se ta delež še precej poveča 6. Pri otrocih disleksičnih staršev so odzivi možganov drugačni že od rojstva naprej. Raziskave s funkcionalno magnetno resonanco so pokazale, da je pri disleksikih funkcijsko povezovanje regij možganov, ki sodelujejo pri branju, precej slabše 7. Drugačni odzivi možganov pri disleksiji so povezani s spremembami v anatomiji primarnega slušnega korteksa oziroma superiotemporalnega korteksa in tudi drugih okcipito-temporalnih, posteriotemporalnih in okcipitalnih regij.
Novejše študije potrjujejo povezavo med tekočim branjem in spremembo v volumnu možganske skorje pri otrocih v razvoju 8, 9. Raziskovalci so ugotovili, da je z učinkovitim branjem povezan upad volumna frontalnega in parietalnega režnja možganske skorje 4. Na podlagi merjenja odzivov možganov pri dojenčku je celo možno napovedati hitrost branja, ki jo bo usvojil v obdobju adolescence 10. Pridobljena nezmožnost branja, aleksija, pa se lahko pojavi pri ljudeh, ki so že brali, vendar so izgubili to sposobnost, bodisi po kapi ali poškodbi področja v možganih, ki je povezano z bralnimi sposobnostmi. Najpogosteje je aleksija posledica infarkta leve posteriorne cerebralne arterije, kar se kaže v tem, da oseba lahko piše, ne more pa brati 11, 12.
V splošnem torej velja, da je intaktna leva hemisfera možganov, še posebno temporoparietalni del, izrednega pomena za dobro bralno sposobnost. Vendar kompleksnost razvoja procesa pridobivanja sposobnosti branja ne temelji le na aktivaciji ustreznih delov skorje, temveč tudi in predvsem na ustrezni komunikaciji med njimi. Raziskave o natančni identifikaciji povezav med anatomijo in funkcijo delov možganov v povezavi z branjem se nadaljujejo in nam bodo v prihodnosti omogočile natančnejši vpogled v dejansko shemo procesa učenja branja.
-
___
-
Vandermosten M., Boets B., Wouters J., Ghesquiere P. A qualitative and quantitative review of diffusion tensor imaging studies in reading and dyslexia. Neuroscience and Biobehavioral Reviews. 2012;36(6):1532-1552. ↩
-
Kremen W. S., Prom-Wormley E., Panizzon M. S., Eyler L. T., Fischl B., Neale M. C., Franc C. E., Lyons M. J., Pacheco J., Perry M. E., Stevens A., Schmitt J. E., Grant M. D., Sediman L. J., Thermenos H. W., Tsuang M. T., Eisen S. A., Dale A. M. Fennema-Notestine C. Genetic and environmental influences on the size of specific brain regions in midlife: the VETSA MRI study. Neuroimage. 2010;49(1):1213-1223. ↩
-
Luby J. L., Barch D. M., Belden A., Gaffrey M. S., Tillman R., Babb C., Nishino T., Suzuki H., Botteron K. N. Maternal support in early childhood predicts larger hippocampal volumes at school age. Proceedings of the National Academy of Sciences of the United States of America. 2012;109(8):2854-2859. ↩
-
Houston S. M., Lebel C., Katzir T., Manis F. R., Kan E., Rodriguez G. R., Sowell E. R. Reading skill and structural brain development. Neuroreport. 2014;25(5):347-352. ↩
-
Vellutino F.R., Fletcher J.M., Snowling M.J., Scanlon D.M. Specific reading disability (dyslexia): what have we learned in the past four decades? Journal of Child Psychology and Psychiatry. 2004;45:2–40. ↩
-
Schumacher J., Hoffmann P., Schmal C., Schulte-Korne G., Nothen M. M. Genetics of dyslexia: the evolving landscape. Journal of Medical Genetics. 2007;44(5):289-297. ↩
-
Boets B., Op de Beeck H. P., Vandermosten M., Scott S. K., Gillebert C. R., Mantini D., Bulthe J., Sunaert S., Wouters J., Ghesquiere P. Intact but less accessible phonetic representations in adults with dyslexia. Science. 2013;342(6163):1251-1254. ↩
-
Sowell E. R., Thompson P. M., Leonard C. M., Welcome S. E., Kan E., Toga A. W. Longitudinal mapping of cortical thickness and brain growth in normal children. Journal of Neuroscience. 2004;24:8223–8231. ↩
-
Squeglia L. M., Jacobus J., Sorg S. F., Jerniga T. L., Tapert ,S. F. Early adolescentcortical thinning is relatedto better neuropsychological performance. Journal of the International Neuropsychological Society. 2013;19:962–970. ↩
-
Lohvansuu K., Hamalainen J. A., Ervast L., Lyytinen H., Leppanen P. H. T. Longitudinal interactions between brain and cognitive measures on reading development from 6 months to 14 years. Neuropsychologia. 2018;108:6-12. ↩
-
Kim E. S., Rising K., Rapcsak S. Z., Beeson P. M. Treatment for Alexia With Agraphia Following Left Ventral Occipito-Temporal Damage: Strengthening Orthographic Representations Common to Reading and Spelling. JSLHR. 2015;58:1521. ↩
-
Ožura A., Horvat Ledinek A., Starovasnik B., Sever A., Vodušek D. B. Aleksija brez agrafije pri tumorski obliki multiple skleroze: prikaz primera. Zdravstveni vestnik. 2009;78:9-12. ↩
Asist. – razisk. dr. Maša Čater, univ. dipl. bioteh.
Laboratorij za genomiko
Inštitut za predklinične vede
Veterinarska fakulteta
Univerza v Ljubljani
Sprejeto: 17.12.2018
Objavljeno: 14.1.2019