Spletna revija za znanstvenike, strokovnjake
in nevroznanstvene navdušence
Naslovnica Članki Intervjuji Mnenja Zdravje Korenine eSinapsa Številke ![]()
Kako so prostorske informacije predstavljene in shranjene v naših možganih?
letnik 2022, številka 23
uvodnik
Kaj je zavest in kje je shranjena?
Maša Čater
članki
Akutni ishemični infarkt hrbtenjače pri zdravih otrocih – kaj lahko pove radiolog?
Katarina Šurlan Popovič, Barbara Šijaković
aktualno
zdravje
Maja Bresjanac
kolofon
letnik 2022, številka 23
Naši možgani nam omogočajo, da obdelujemo informacije iz okolja ter na njihovi podlagi oblikujemo miselne predstave zunanjega sveta. Pomemben vidik naše predstave zunanjega sveta so prostorske informacije, saj vse, kar nas obdaja, zavzema mesto oziroma položaj v prostoru. Prostorske informacije pa niso pomembne zgolj za naše zaznavanje, temveč imajo ključno vlogo tudi pri usmerjanju naših gibov in vedênja. Vsak gib je skrbno preračunan na podlagi položaja našega telesa ter njegovega odnosa do objektov v prostoru. Poraja pa se vprašanje, na kakšne načine naši možgani zabeležijo, shranijo in uporabijo tovrstne prostorske informacije. Razumevanje prostorskih predstav ter njihovih nevrofizioloških mehanizmov ostaja pomemben izziv kognitivne nevroznanosti.
Nevrofiziologija prostorskega delovnega spomina
Sposobnost kratkoročnega shranjevanja prostorskih informacij, ki so pomembne za naše vedênje, omogoča prostorski delovni spomin. Prostorski delovni spomin se pogosto preučuje s pomočjo vedenjskih preizkušenj, kjer udeležencu za kratek čas predstavimo vidni dražljaj, ki mu sledi nekaj sekundni zamik brez prikaza dražljaja (Slika 1) 1 2 3. Naloga udeleženca je zapomniti si točen položaj dražljaja, ki ga mora po zamiku priklicati z izvedbo giba na ta položaj, kot na primer s premikom roke ali pogleda. Predpostavlja se, da takšna naloga zahteva kratkoročno shranjevanje prostorskih informacij za uspešno izvedbo naloge, saj v času zamika naloge senzorični dražjaj ni več neposredno dostopen preko naših čutov.
Tovrstno nalogo so uporabili v številnih raziskavah za preučevanje nevrofizioloških mehanizmov prostorskega delovnega spomina s pomočjo elektrofizioloških meritev na živalih 4 5 6 ali nevroslikovnimi metodami (npr. pozitronska emisijska tomografija – PET, funkcijska magnetna resonanca – fMR) na ljudeh 7 8 9. Konsistenten izsledek teh raziskav je povišana aktivnost v širokem razponu možganskih področij v frontalnem in parietalnem režnju ter posteriornih senzoričnih regijah, ki je vzdrževana v času zamika naloge (Slika 2). Predpostavlja se, da tovrstna možganska aktivnost odraža aktivno vzdrževanje prostorskih informacij ter tako predstavlja nevrofiziološki korelat prostorskega delovnega spomina 10.
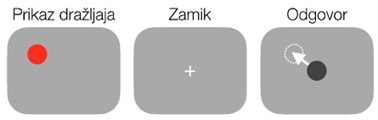
Slika 1: Primer vedenjske preizkušnje prostorskega delovnega spomina. Udeležencem je za kratek čas prikazan vidni dražljaj, ki izgine v času nekaj sekundnega zamika. Naloga udeleženca je zapomniti si točen položaj dražljaja in ga po zamiku priklicati s pomočjo premika roke ali pogleda na ta položaj.
Možganski koordinatni sistem
Ključno opažanje, ki kaže na prisotnost specifičnih prostorskih predstav v vzdrževani aktivnosti posameznih živčnih celic v času zamika naloge prostorskega delovnega spomina, je selektivna občutljivost teh celic na specifično lokacijo senzoričnega dražljaja 4 5 6. Tovrstna selektivna aktivnost se kaže le, kadar je dražljaj, ki si ga je potrebno zapomniti, prikazan v določenem območju ali smeri vidnega polja. Na ta način se specifične prostorske informacije beležijo v različnih vzorcih aktivnosti posameznih živčnih celic. Številne študije so pokazale, da so živčni odzivi na prostorske dražljaje topografsko organizirani, kar pomeni, da se pri beleženju prostorskih informacij v možganih ohranijo prostorska razmerja, kot so prikazana v vidnem polju (npr. sosednja položaja v vidnem polju sprožita aktivnost v sosednjih živčnih celicah) 11. Kljub dobro opisanim značilnostim živčnih odzivov na prostorske dražljaje pa ostaja vprašanje, kako bi lahko te prostorske predstave opisali v smislu znanih metričnih sistemov.
Za določanje položaja dražljaja morajo naši možgani oblikovati nekakšen miselni koordinatni sistem, ki se v literaturi pogosto opisuje kot referenčni okvir. V matematičnem smislu referenčni okvir zahteva referenčno točko (izhodišče) in referenčne smeri (koordinatne osi, npr. horizontalna in vertikalna os v kartezičnem koordinatnem sistemu ali kot in amplituda v polarnem koordinatnem sistemu), pri čemer lahko vsak položaj določimo z nizom koordinat 12. Raziskave so pokazale obstoj različnih referenčnih okvirjev glede na referenčno točko 13 14. Na primer, lokacija je lahko zabeležena relativno na naš pogled oziroma telo (egocentrični referenčni okvir) ali glede na druge predmete v vidnem polju (alocentrični referenčni okvir). Manj pa je znanega o vrsti referenčnih osi pri beleženju prostorskih informacij. Na podlagi opažanj, da se posamezne živčne celice selektivno odzivajo na kot in amplitudo zapomnjenega položaja dražljaja, se predpostavlja polarno kodiranje prostorskih informacij v procesih delovnega spomina 4 6 15.
Strategije shranjevanja in vzdrževanja prostorskih informacij
Ne glede na obliko prostorskih predstav se domneva, da za shranjevanje in vzdrževanje prostorskih informacij uporabljamo različne miselne strategije. V kontekstu preizkušnje prostorskega delovnega spomina so številni avtorji predlagali, da je položaj dražljaja v času zamika naloge lahko shranjen bodisi kot pretekla senzorična informacija o zaznanem dražljaju (retrospektivno senzorično kodiranje), bodisi kot motorični načrt, pripravljen za izvedbo odziva na ta dražljaj (prospektivno motorično kodiranje) 1 3 10. Na ravni retrospektivnega senzoričnega kodiranja se predpostavlja, da se prostorske predstave vzdržujejo s pomočjo usmerjanja pozornosti na lokacijo senzoričnega dražljaja, tudi ko dražljaj več ni prikazan. Po drugi strani pa naši odzivi zahtevajo pretvorbo senzoričnih predstav v motorične načrte, ki usmerjajo naše gibe. Vzdrževanje prostorskih predstav s pomočjo prospektivnega motoričnega kodiranja bi potemtakem pomenilo zgolj reaktivacijo motoričnih programov, ki bi jih lahko uporabili kot strategijo kodiranja tudi v primeru, ko odziv ni dejansko izveden (npr. zapomnili bi si premik oziroma pot pogleda ali roke do tarčnega dražljaja).
V podporo predpostavki o retrospektivnem senzoričnem in prospektivnem motoričnem kodiranju so številna opažanja, da se posamezne živčne celice odzivajo selektivno na senzorične prostorske informacije ali smer motoričnega odziva 4 6. fMR-raziskave na ljudeh so pokazale, da je možno vplivati na vrsto uporabljene strategije z manipulacijo zahtev vedenjske preizkušnje, ki se odraža v spremembah možganske aktivnosti 2 3. V izvirni raziskavi Purg idr. 3 smo s pomočjo fMR meritev med izvedbo naloge prostorskega delovnega spomina pokazali, da se retrospektivno senzorično kodiranje povezuje s povišano aktivnostjo in funkcijsko povezanostjo v okcipitalnih in parietalnih področjih, povezanih z vidnimi in pozornostnimi procesi. Po drugi strani pa je bilo prospektivno motorično kodiranje povezano s povišano aktivnostjo in funkcijsko povezanostjo v somatosenzoričnih, premotoričnih in motoričnih možganskih področjih.
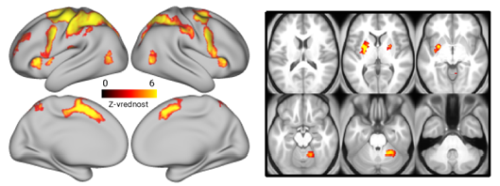
Slika 2: Možganska področja, ki kažejo povišano aktivnost v času zamika naloge prostorskega delovnega spomina. Prilagojeno po Purg idr. 3.
Zaključki
Obdelava in shranjevanje prostorskih informacij v možganih je pomembna tema v raziskovanju na področju kognitivne nevroznanosti, ki že sedaj obsega precejšen razpon literature. Kljub dobro opisanim vedenjskim in nevrofiziološkim mehanizmom kratkoročnega shranjevanja prostorskih informacij pa ostaja še precej odprtih vprašanj. Izziv prihodnjih raziskav je preučiti specifične oblike prostorskih predstav in njihove pretvorbe za uporabo v različnih kognitivnih procesih. Pomembna smer raziskovanja so tudi individualne razlike v procesih prostorskega delovnega spomina, ki bi lahko razkrile, kako se ljudje razlikujemo v zaznavi, shranjevanju in uporabi prostorskih informacij.
-
___
-
Funahashi, S. (2013). Space representation in the prefrontal cortex. Progress in Neurobiology, 103, 131-155. doi: 10.1016/j.pneurobio.2012.04.002 ↩
-
Curtis, C. E., Rao, V. Y., in D’Esposito, M. (2004). Maintenance of spatial and motor codes during oculomotor delayed response tasks. Journal of Neuroscience, 24(16), 3944-3952. doi: 10.1523/ JNEUROSCI.5640-03.2004 ↩
-
Purg, N., Starc, M., Slana Ozimič, A., Kraljič, A., Matkovič, A., in Repovš, G. (2022). Neural Evidence for Different Types of Position Coding Strategies in Spatial Working Memory. Frontiers in Human Neuroscience, 16:821545. doi: 10.3389/fnhum.2022.821545 ↩
-
Fuster, J. M., in Alexander, G. E. (1971). Neuron activity related to short-term memory. Science, 173(3997), 652-654. doi: 10.1126/science.173.3997.652 ↩
-
Fuster, J. M. (1973). Unit activity in prefrontal cortex during delayed-response performance: Neuronal correlates of transient memory. Journal of Neurophysiology, 36(1), 61-78. doi: 10.1152/jn.1973.36.1.61 ↩
-
Funahashi, S., Bruce, C. J., in Goldman-Rakic, P. S. (1989). Mnemonic coding of visual space in the monkey’s dorsolateral prefrontal cortex. Journal of Neurophysiology, 61(2), 331-349. doi: 10.1152/ jn.1989.61.2.331 ↩
-
Sweeney, J. A., Mintun, M. A., Kwee, S., Wiseman, M. B., Brown, D. L., Rosenberg, D. R., in Carl, J. R. (1996). Positron emission tomography study of voluntary saccadic eye movements and spatial working memory. Journal of Neurophysiology, 75(1), 454-468. doi: 10.1152/jn.1996.75.1.454 ↩
-
Zarahn, E., Aquirre, G. K., in D’Esposito, M. (1999). Temporal isolation of the neural correlates of spatial mnemonic processing with fMRI. Brain Research, Cognitive Brain Research, 7(3), 255-268. doi: 10.1016/ s0926-6410(98)00029-9 ↩
-
Brown, M. R. G., DeSouza, J. F. X., Goltz, H. C., Ford, K., Menon, R. S., Goodale, M. A., in Everling, S. (2004). Comparison of memory- and visually-guided saccades using event-related fMRI. Journal of Neurophysiology, 91(2), 873-889. doi: 10.1152/jn.00382.2003 ↩
-
D’Esposito, M., in Postle, B. R. (2015). The cognitive neuroscience of working memory. Annual Review of Psychology, 66, 115-142. doi: 10.1146/annurev-psych-010814-015031 ↩
-
Silver, M. A., in Kastner, S. (2009). Topographic maps in human frontal and parietal cortex. Trends in Cognitive Science, 13(11), 488-495. doi: 10.1016/j.tics.2009.08.005 ↩
-
Wang, R. F. (2012). Theories of spatial representations and reference frames: what can configuration errors tell us? Psychonomic Bulletin & Review, 19(4), 575-587. doi: 10.3758/s13423-012-0258-2 ↩
-
Burgess, N. (2006). Spatial memory: how egocentric and allocentric combine. Trends in Cognitive Science, 10(12), 551-557. doi: 10.1016/j.tics.2006.10.005 ↩
-
Tatler, B. W., in Land, M. F. (2011). Vision and the representation of the surroundings in spatial memory. Philosophical Transactions of the Royal Society B: Biological Sciences, 366(1564), 596-610. doi: 10.1098/rstb.2010.0188 ↩
-
Swisher, J. D., Halko, M. A., Merabet, L. B., McMains, S. A., in Somers, D. C. (2007). Visual topography of human intraparietal sulcus. Journal of Neuroscience, 27(20), 5326-5337. doi: 10.1523/ JNEUROSCI.0991-07.2007 ↩
asist. dr. Nina Purg
Oddelek za psihologijo
Filozofska fakulteta
Univerza v Ljubljani
Sprejeto: 6.1.2023
Objavljeno: 18.1.2023