Spletna revija za znanstvenike, strokovnjake
in nevroznanstvene navdušence
Naslovnica Članki Intervjuji Mnenja Zdravje Korenine eSinapsa Številke ![]()
O lososih, funkcijski magnetni resonanci in izzivih statističnega testiranja
članki
eSinapsa, 2011-1
Zvezdan Pirtošek
Eksoskeleti – inteligentne bionske naprave
Marko Munih
O aktualnih dilemah draženja globokih možganskih struktur pri obsesivno - kompulzivni motnji
Nadja Jarc
Sledite svojo srečo ... z iPhone
Urban Kordeš
eSinapsa, 2011-2
Renata Salecl
Gašper Tkačik
Astrociti – spregledane zvezde nevrobiologije
Marko Kreft, Robert Zorec
Sašo Dolenc
Meditacija - malo truda, veliko koristi
Luka Dimic
eSinapsa, 2011-3
Mara Bresjanac
Martina Starc
Rok Berlot
Varnost uporabe generičnih protiepileptičnih zdravil
Mojca Kržan, Matevž Kržan
Možgani, računalniki - nekaj vmes
Miha Pelko
eSinapsa, 2012-4
Ali so moški in ženski možgani različni?
Gregor Majdič
O kognitivnih motnjah pri bolnikih s Parkinsonovo boleznijo
Dejan Georgiev
Akutno možgansko kap lahko uspešno zdravimo
Nina Vujasinovič, Bojana Žvan
Vloga nevropsihološke diagnostike pri odkrivanju zgodnjih znakov alzheimerjeve bolezni
Simon Brezovar
eSinapsa, 2013-5
Subarahnoidna krvavitev zaradi tromboze venskih sinusov
Mateja Repar, Anita Resman Gašperčič
Srečanje dveh velikanov: možganov in imunskega sistema
Matej Markota
Novo odkritje na področju sporadičnih prionskih bolezni
Jana Jerše, Nadja Jarc
Učinek placeba brez lažnih zdravil in zavajanja
Mara Bresjanac
eSinapsa, 2013-6
Odstranjevanje možganskih tumorjev pri budnem bolniku
Andrej Vranič, Jasmina Markovič, Blaž Koritnik
Zmedena bolnica, ki nič ne vidi ali PRES
Manja Hribar, Vid Zgonc
Manja Hribar
Sistemska skleroza in ishemična možganska kap - vzročna povezanost ali le koincidenca?
Mateja Repar, Janja Pretnar Oblak
Netravmatska lokalizirana konveksitetna subarahnoidna krvavitev
Mateja Repar, Fajko F. Bajrović
Klemen Grabljevec
Z omejevanjem spodbujajoča terapija pri bolnikih po nezgodni možganski poškodbi
Dejana Zajc, Klemen Grabljevec
eSinapsa, 2014-7
Možgani v mreži navezanosti, ki nas zaznamuje
Barbara Horvat
Vpliv senzoričnega dotoka na uglasitev možganskih povezav
Peter Gradišnik
Človeški konektom ali kakšne so zveze v naših možganih
Blaž Koritnik
Niko Lah
Torkove delavnice za osnovnošolce
Mateja Drolec Novak, Vid V. Vodušek
Da ne pozabim! Tehnike za pomladitev spomina
Klara Tostovršnik, Hana Hawlina
Površina socialne nevroznanosti
Manuel Kuran
Clarity - bistri možgani Karla Deisserotha
Gregor Belušič
Barbara Gnidovec Stražišar
Bojana Žvan
Nevroplastičnost po možganski kapi
Marjan Zaletel
Klinično psihološka obravnava pacientov po možganski kapi in podpora pri vračanju na delovno mesto
Barbara Starovasnik Žagavec
Možgani: organ, s katerim ljubimo
Andraž Matkovič
Marija Šoštarič Podlesnik
Gibalno-kognitivna vadba: praktična delavnica
Mitja Gerževič, Marina Dobnik
Anton Grad
Nevrologija, imunologija, psihiatrija …
Bojan Rojc
Andraž Stožer, Janez Bregant
Dominika Novak Pihler
Možganska kap – »kako ostati v omrežju?«
Nina Ozimic
Klara Tostovršnik
eSinapsa, 2014-8
Znotrajžilno zdravljenje možganskih anevrizem
Tamara Gorjanc, Dimitrij Lovrič
Obravnava hladnih možganskih anevrizem
Bojana Žvan, Janja Pretnar Oblak
Ali deklice z Rettovim sindromom govorijo z očmi?
Anka Slana, Urška Slana
Progresivna multifokalna encefalopatija
Urša Zabret, Katarina Šurlan Popovič
Ne ubijaj – poskusi na živalih
Martina Perše
Poizkusi na živalih - za in proti
Simon Horvat
eSinapsa, 2015-9
Kako deluje navigacijski sistem v naših možganih
Simon Brezovar
Vsakodnevno delo slepe osebe / s slepo osebo
Denis Kamnar
Uroš Marušič
Manca Tekavčič Pompe
Toni Pustovrh
Marko Hawlina
Od svetlobe do podobe ali kako vidijo svet naši možgani
Simon Brezovar
Janja Hrastovšek
Zala Kurinčič
Pogledi na mejno osebnostno motnjo
Jerica Radež, Peter Kapš
Uvid kot socialno psihološki fenomen
Vid Vodušek
Uvod v vidno-prostorske funkcije s praktičnimi primeri
Ana Bujišić, Sanja Roškar
eSinapsa, 2015-10
Difuzijsko magnetnoresonančno slikanje
Rok Berlot
Katja Pavšič
Radiološko izolirani sindrom - ali ga moramo poznati?
Matej Vouk, Katarina Šurlan Popovič
Kako izgledajo možgani, ki govorijo več jezikov?
Gašper Zupan
Nov pristop v rehabilitaciji - terapija s pomočjo psa
Mateja Drljepan
Pogled v maternico z magnetnoresonančno preiskavo
Taja Jordan, Tina Vipotnik Vesnaver
Saša Zorjan
Saša Zorjan
Nevroestetika: ko nevroznanost obišče galerijo
Anja Voljavec, Hana Hawlina, Nika Vrabič
Ali so psihogeni neepileptični napadi res psihogeni?
Saška Vipotnik, Gal Granda
Kako nam lahko glasna glasba »vzame« sluh in povzroči tinitus
Nejc Steiner, Saba Battelino
eSinapsa, 2016-11
Mara Bresjanac
Kako ultrazvok odpira pot v možgane
Kaja Kolmančič
Kako je epigenetika spremenila nevroznanost
Metka Ravnik Glavač
Ondinino prekletstvo ali sindrom prirojene centralne hipoventilacije
Katja Pavšič, Barbara Gnidovec Stražišar, Janja Pretnar Oblak, Fajko F. Bajrović
Zika virus in magnetnoresonančna diagnostika nepravilnosti osrednjega živčevja pri plodu
Rok Banko, Tina Vipotnik Vesnaver
Motnje ravnotežja otrok in odraslih
Nejc Steiner, Saba Battelino
eSinapsa, 2016-12
Vloga magnetnoresonančne spektroskopije pri obravnavi možganskih tumorjev
Gašper Zupan, Katarina Šurlan Popovič
Tiskanje tridimenzionalnih modelov v medicini
Andrej Vovk
Aleš Oblak
Kevin Klarič
Sinestezija: umetnica, ki ne želi odrasti
Tisa Frelih
Računska psihiatrija: od nevroznanosti do klinike
Nastja Tomat
Kognitivni nadzor: od vsakdanjega življenja do bolezni
Vida Ana Politakis
eSinapsa, 2017-13
Internet: nadgradnja ali nadomestek uma?
Matej Perovnik
Vloga črevesnega mikrobioma pri odzivu na stres
Vesna van Midden
Stres pušča posledice tako na človeškem kot živalskem organizmu
Jasmina Kerčmar
Prikaz normalne anatomije in bolezenskih stanj obraznega živca z magnetno resonanco
Rok Banko, Matej Vrabec
Psihedelična izkušnja in njen zdravilni potencial
Anja Cehnar, Jona Basle
Vpliv hiperglikemije na delovanje možganov
Jasna Šuput Omladič, Simona Klemenčič
Nevrofibromatoza: napredujoče obolenje centralnega in perifernega živčevja
Nejc Steiner, Saba Battelino
Fenomen žrtvenega jagnja v dobi interneta
Dolores Trol
Tesnoba staršev in strategije spoprijemanja, ko pri otroku na novo odkrijejo epilepsijo
Daša Kocjančič, Petra Lešnik Musek, Vesna Krkoč, David Gosar
eSinapsa, 2017-14
Zakaj ne zapeljem s ceste, ko kihnem?
Anka Slana Ozimič, Grega Repovš
Možgani pod stresom: od celic do duševnih motenj
Nastja Tomat
Nobelova nagrada za odkritje molekularnih mehanizmov nadzora cirkadianih ritmov
Leja Dolenc Grošelj
Na sledi prvi vzročni terapiji Huntingtonove bolezni
Danaja Metul
Razlike med spoloma pri Parkinsonovi bolezni
Kaja Kolmančič
eSinapsa, 2018-15
Susceptibilno poudarjeno magnetnoresonančno slikanje pri bolniku z ALS
Alja Vičič, Jernej Avsenik, Rok Berlot
Sara Fabjan
Reverzibilni cerebralni vazokonstrikcijski sindrom – pot do diagnoze
Maja Cimperšek, Katarina Šurlan Popovič
Liam Korošec Hudnik
Kognitivno funkcioniranje pri izgorelosti
Marina Horvat
eSinapsa, 2019-16
Maša Čater
Saša Koprivec
Infekcije osrednjega živčnega sistema s flavivirusi
Maja Potokar
Raziskava: Kako depresija vpliva na kognitivne sposobnosti?
Vida Ana Politakis
Razvoj depresije pri otrocih z vidika navezovalnega vedenja
Neža Grgurevič
Sonja Prpar Mihevc
Umetno inteligentna nevroznanost: srečanje nevronskih mrež in možganske fiziologije
Kristijan Armeni
Čebelji strup pri preventivi nevrodegenerativnih bolezni in priložnost za klinično prakso
Matjaž Deželak
eSinapsa, 2019-17
IgG4+ – skupni imenovalec diagnoz iz preteklosti
Cene Jerele, Katarina Šurlan Popovič
Nov molekulski mehanizem delovanja ketamina v astrocitih
Matjaž Stenovec
Praktični pristop k obravnavi utrujenosti in motenj spanja pri bolnikih z multiplo sklerozo
Nik Krajnc, Leja Dolenc Grošelj
Jure Pešak
eSinapsa, 2020-18
Bolezni spektra anti-MOG pri odraslih
Nik Krajnc
Samomor pod lupo nevroznanosti
Alina Holnthaner
eSinapsa, 2020-19
Ob mednarodnem dnevu znakovnih jezikov
Anka Slana Ozimič
Teorija obetov: kako sprejemamo tvegane odločitve
Nastja Tomat
Sara Fabjan
Matjaž Deželak
Nina Stanojević, Uroš Kovačič
Od človeških nevronov do možganskih organoidov – nova obzorja v nevroznanosti
Vesna M. van Midden
Splošna umetna inteligenca ali statistične jezikovne papige?
Kristijan Armeni
Zunajcelični vezikli kot prenašalci zdravilnih učinkovin preko krvno-možganske prepreke
Saša Koprivec
Matjaž Deželak
eSinapsa, 2021-20
Migrena: starodavna bolezen, sodobni pristopi k zdravljenju
Eva Koban, Lina Savšek
Zgodnji razvoj socialnega vedenja
Vesna Jug
Nastja Tomat
Mikrosplet: povezovanje preko mikrobioma
Tina Tinkara Peternelj
Stimulacija možganov kot način zdravljenja depresije
Saša Kocijančič Azzaoui
eSinapsa, 2021-21
eSinapsa, 2022-22
Sodobni vidiki motenj hranjenja
Karin Sernec
Ples in gibalni dialog z malčki
Neva Kralj
Atul Gawande
Jezikovna funkcija pri Alzheimerjevi bolezni
Gašper Tonin
Dostava terapevtikov preko krvno-možganske pregrade
Matjaž Deželak
eSinapsa, 2022-23
Akutni ishemični infarkt hrbtenjače pri zdravih otrocih – kaj lahko pove radiolog?
Katarina Šurlan Popovič, Barbara Šijaković
eSinapsa, 2023-24
Možganska omrežja pri nevrodegenerativnih boleznih
Tomaž Rus, Matej Perovnik
Morske živali kot navdih za nevroznanstvenike: morski konjiček, morski zajček in klobučnjak
Tina Bregant
Metoda Feldenkrais: gibanje in nevroplastičnost
Mateja Pate
Etično naravnana animalna nevroznanost
Maša Čater
Helena Motaln, Boris Rogelj
eSinapsa, 2023-25
Urban Košak, Damijan Knez, Anže Meden, Simon Žakelj, Jurij Trontelj, Jure Stojan, Maja Zakošek Pipan, Kinga Sałat idr.
eSinapsa, 2024-26
Naravno okolje kot vir zdravja in blagostanja
Karin Križman, Grega Repovš, Gaja Zager Kocjan, Gregor Geršak
Katja Peganc Nunčič, Damjan Osredkar
Leta 2009 je na konferenci Organization for Human Brain Mapping precej ironičnega navdušenja pritegnil poster z naslovom “Neural correlates of interspecies perspective taking in the post-mortem Atlantic Salmon: An argument for multiple comparisons correction”1, v katerem so avtorji s pomočjo funkcijske magnetne resonance (fMR) identificirali področje možganov, ki je bilo v mrtvem lososu “aktivno” pri izvedbi preizkušnje prepoznavanja čustvenih izrazov na slikah človeških obrazov. Rezultati so bili seveda nesmiselni, avtorji pa so želeli z njimi opozoriti na nekatere izzive veljavne izvedbe fMR študij.
 Poster je precej zanimanja požel tudi zunaj znanstvene skupnosti, ki uporablja slikovne tehnike preučevanja delovanja možganov in duševnosti ter postal internetni meme na katerega radi opozorijo tudi predstavniki laične javnosti, ko pogovor nanese na fMR. Rezultati, ki jih omogoča, so tako ali tako lari-fari, je pogost zaključek. Po počasnih začetkih se - ob pridobitvi raziskovanju posvečenega tomografa na Medicinski fakulteti Univerze v Ljubljani - na pot uporabe fMR v okviru kognitivne nevroznanosti odločneje podajamo tudi v Sloveniji. Ob tej priložnosti je smiselno nasloviti tudi kritike, ki jih bodisi neposredno podaja bodisi posredno odpira omenjeni prispevek.
Poster je precej zanimanja požel tudi zunaj znanstvene skupnosti, ki uporablja slikovne tehnike preučevanja delovanja možganov in duševnosti ter postal internetni meme na katerega radi opozorijo tudi predstavniki laične javnosti, ko pogovor nanese na fMR. Rezultati, ki jih omogoča, so tako ali tako lari-fari, je pogost zaključek. Po počasnih začetkih se - ob pridobitvi raziskovanju posvečenega tomografa na Medicinski fakulteti Univerze v Ljubljani - na pot uporabe fMR v okviru kognitivne nevroznanosti odločneje podajamo tudi v Sloveniji. Ob tej priložnosti je smiselno nasloviti tudi kritike, ki jih bodisi neposredno podaja bodisi posredno odpira omenjeni prispevek.
fMR in problem multiplih primerjav
V svojem prispevku se avtorji osredotočajo predvsem na t.i. problem multiplih primerjav. Za njegovo razumevanje je treba podati kratke osnove uporabe fMR (nekoliko širšo, a vseeno dostopno predstavitev lahko najdete v več virih, npr. 1 ali 2). Kadar želimo ugotoviti katera področja možganov so aktivna pri izvedbi določene naloge, udeležence v raziskavi prosimo, da omenjeno nalogo izvajajo v točno opredeljenem času, med katerim hkrati s pomočjo fMR zajemamo niz slik možganov, na katerih je intenzivnost signala odvisna od stopnje oksigeniranosti hemoglobina v krvi. Tovrsten niz slik nam omogoča sledenje t.i. BOLD (Blood Oxygenation Level Dependent) signala, za katerega vemo, da se tekom izvedbe naloge spreminja. Kadar se aktivnost v določenem predelu možganov poveča, se z njo - zaradi povečanih energetskih potreb - poveča tudi poraba kisika. Krvno-žilni sistem se na povečano porabo kisika odzove z večjim pretokom krvi v relevantnem področju. Povečanje pretoka je venomer večje od potreb, zato pride do prehodnega povečanja količine oksigeniranega hemoglobina, kar se odraža v zajetem signalu. Ker zaradi podrobnega časovnega nadzora vemo, kdaj je prišlo do izvedbe naloge, lahko napovemo do kakšne spremembe signala bi moralo priti v področjih možganov, ki so vpleteni v izvajanje naloge. Tovrstna področja prepoznamo s pomočjo multiple regresije tako, da za vsako volumensko enoto slike (voksel) preverimo, ali lahko z napovedanim signalom pojasnimo pomemben del opazovanega signala. Če predvideni signal pomembno napove opazovani signal, lahko zaključimo, da je področje, od koder izvira opazovan signal, udeleženo v izvedbi naloge.
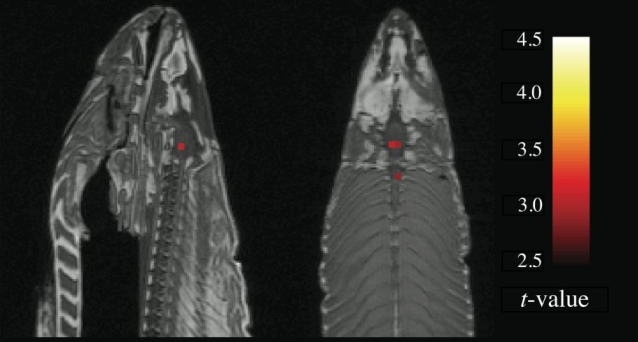
Losos, pri katerem je statistična analiza brez ustreznih popravkov pokazala 16 aktivnih vokslov od 8064 na ravni p < ,001 (Vir: prefrontal.org).
Ko presojamo pomembnost napovedi, se naslonimo na orodja statističnega testiranja. Logika statističnega testiranja je, poenostavljeno povedano, naslednja. S pomočjo statističnih testov lahko ocenimo verjetnost, da bi do opazovanega ali večjega učinka lahko prišlo po naključju. Če je ta verjetnost (največkrat označena kot vrednost p) zelo majhna, potem si upamo zaključiti, da opazovana velikost učinka ni naključna, temveč odraža dejanski pojav. Najpogosteje uporabljena mejna p vrednost v statističnem testiranju je 0,05. Z drugimi besedami, če je verjetnost, da je opazovani rezultat naključen manj kot petodstotna, smo pripravljeni zavreči možnost, da opazujemo le naključje in sprejmemo trditev, da je prišlo do statistično pomembnega vpliva preučevanega dejavnika. V primeru fMR bi sprejeli trditev, da sprememba ob izvajanju podane naloge v opazovanem BOLD-signalu ne odraža le šuma ali z nalogo nepovezanih aktivnosti, temveč gre pri tem za možgansko aktivnost.
Problem, ki ga poudarjajo avtorji je, da v omenjenem postopku statističnega testa ne izvedemo le enkrat, temveč ga izvajamo za vsak voksel posebej. Ker so verjetnosti naključnega preseganja zastavljenega merila (načeloma) neodvisne, se z večanjem števila testov povečuje tudi verjetnost, da bo mejna vrednost v kateremkoli izmed njih presežena po naključju. Za razumevanje problema bo na tem mestu morda dobrodošla analogija. Negativen učinek ionizirajočega sevanja ob rentgentskem slikanju je razmeroma dobro znan. Verjetnost, da bi zaradi posamičnega slikanja prišlo do razvoja rakavega obolenja je izredno majhna, s pogosto izpostavljenostjo pa se le-ta veča, zato se nepotrebnemu slikanju izogibamo. Enako je pri statističnem testiranju: verjetnost preseganja mejne vrednosti v posamičnem testu je razmeroma majhna, z večanjem števila testov pa se ta verjetnost povečuje. Pri uporabi mejne vrednosti 0.05, lahko pričakujemo, da bo pri izvedbi stotih testov rezultat po naključju presegel mejno vrednost pri petih izmed njih. Preneseno na fMR lahko pri testiranju 60.000 vokslov pričakujemo preseganje mejne vrednosti p < 0,05 pri 3.000 izmed njih. Na ta način lahko ugotovimo tudi pomembno odzivnost možganov mrtvega lososa v nalogi presojanja emocij.
Preučevanje korelatov duševne aktivnosti s pomočjo fMR seveda ni edino področje, ki je izpostavljeno problemu multiplih primerjav. V preiskovanju povezav med psihološkimi potezami in različnimi vedenjskimi lastnostmi posameznikov so raziskovalci ob odsotnosti uporabe ustreznih korektivnih postopkov lahko hitro soočeni z več deset do več sto korelacijami in posledično napačnim zaključkom. Podobno lahko v genetskih študijah opazujemo povezave med stotinami različnih genov ter osebnostnimi lastnostmi, inteligentnostjo, spolno usmerjenostjo ali tveganjem za različne bolezni in ni čudno, da časopisje vsake toliko poroča o odkritju gena za homoseksualnost, debelost ali inteligentnost, ki zaradi odsotnosti dodatnih potrditev kmalu pade v pozabo.
Korekcija kriterija za naslavljanje problema multiplih povezav
Eden izmed najbolj preprostih orodij v boju proti neveljavnim zaključkom zaradi problema multiplih povezav je uporaba ustreznih korekcijskih metod in postopkov. Najbolj preprost in strog je t.i. Bonfferonijev popravek. Ta temelji na železni logiki. Če želimo možnost, da bi katerikoli statistični test po naključju presegel mejno vrednost, ohraniti na želeni meji, potem je treba želeno mejno vrednost deliti s skupnim številom statističnih testov. Pri 100 testih je nova mejna vrednost 0,0005, pri 60,000 testih 0,00000083.
Medtem ko se z uporabo Bonfferonijevega popravka učinkovito izognemo napaki napačnih prepoznav učinka, smo pri njegovi uporabi soočeni s tako strogim merilom, da mu težko ustreza tudi najbolj robusten učinek - izpostavimo se velikemu deležu napačnih zavrnitev učinka. V fMR praksi se je tudi zato uveljavilo več alternativnih postopkov korekcije za multiple primerjave. Ob predpostavki, da je šum naključno razporejen preko zajetih slik možganov, lahko nekoliko strožje merilo kombiniramo z zahtevo, da se mora učinek pojaviti v več zveznih vokslih4. Koliko vokslov mora presegati mejno vrednost, lahko izračunamo analitično ali s pomočjo simulacij monte-carlo. Uporabimo lahko tudi nekoliko drugačen pristop h kontroli napačnih prepoznav, kot ga predstavlja na primer v izvirnem prispevku omenjeni FDR5 (False Discovery Rate), ali pa uporabimo moderne tehnike ponovnega vzorčenja6 (resampling, bootstrapping).
Naštete metode so pogosto del programskih paketov za fMR analize, ali pa so zajete v samostojnih programih. Ob njihovi smiselni uporabi in ozaveščenosti recenzentov znanstvenih revij je prepoznava aktivnosti v mrtvih lososovih možganih že več let le neposrečena parodija začetkov nove metode preučevanja možganov in njenih neveščih uporabnikov.
Povezave z vedenjskimi rezultati
Poleg neustreznega nadzora nad multiplimi povezavami gre študiji z lososom očitat še eno pomembno pomanjkljivost, ki je morda namenoma botrovala izbiri mrtvega udeleženca - v študiji je umanjkal vedenjski nadzor izvedbe naloge. Da lahko opazovani BOLD-signal povezujemo z izvedbo naloge, moramo biti prepričani, da je udeleženec nalogo zares izvajal. V dejanskih fMR študijah od udeležencev praviloma zahtevamo merljiv vedenjski odziv, ki nam omogoča oceniti, ali in kako dobro je udeleženec izvajal nalogo.
Spremljanje vedenjskih odzivov nam ne pomaga le, da se ubranimo pred napačnimi zaključki, temveč nam omogoča tudi večjo moč pri prepoznavanju relevantnih možganskih področij, saj smo lahko bolj natančni pri napovedi možganske aktivnosti, ter postavljanju podrobnejših vprašanj, na primer, kakšen vzorec možganske aktivnosti loči med uspešnim in neuspešnim kodiranjem informacij v spomin. In kar je morda najpomembnejše: ugotavljanje pomembne povezave med aktivnostjo določenega področja možganov in stopnje pravilnosti izvedbe naloge nam daje najbolj neposreden dokaz za relevantnost prepoznanega področja možganov za izvedbo izbrane naloge.
Multivariatna analiza vzorcev
Verjetno je najbolj učinkovito spoprijemanje z izzivom multiplih primerjav - izogibanje multiplim primerjavam. To je možno na dva načina. Prva možnost sloni na jasno zastavljeni teoriji in specifični domnevi, ki nam omogočata, da statistični test izvedemo le za signal iz jasno opredeljenega področja možganov namesto na tisočih vokslih preko celotnih možganov. Druga možnost pa temelji na uporabi skupka analitičnih metod s skupnim nazivom multivariatna analiza vzorca (ang. Multivariate Pattern Analysis - MVPA).
V okviru multivariatne analize vzorcev si ne zastavimo vprašanja, katera področja možganov pomembno korelirajo s predvidenim BOLD signalom, temveč skušamo ugotoviti, kakšen vzorec aktivnosti najbolje napoveduje, katero nalogo je izvajal (ali kakšnemu dražljaju je bil izpostavljen) udeleženec. Za prepoznavanje vzorcev lahko uporabimo katerega od več možnih postopkov, kot so na primer SVM (Support Vector Machine), ali ANN (Artificial Neural Networks), ki nam povedo, kakšen vzorec aktivnosti se povezuje s posamično nalogo, kot tudi, katera informacija (aktivnost v katerih vokslih) je najbolj diagnostična.
Z uporabo MVPA se izognemo neobvladljivemu številu statističnih testov, saj moramo preveriti le, ali prepoznan vzorec omogoča boljšo napoved kot bi je bili sposobni le po naključju, z metanjem kovanca. Hkrati rezultat podaja zelo uporabno dodatno informacijo in odpira popolnoma nove možnosti uporabe fMR. Ob vrednotenju prepoznanega vzorca ne izvemo le, ali je vzorec pomembno boljši od naključja, temveč tudi, kako dober je, v neposredno razumljivi in uporabni obliki. Vzorec, ki v izbiri med dvema možnostma podaja pravilno napoved v 55 % primerov, je, čeprav boljši od kovanca, vsekakor manj uporaben kot vzorec, ki pravilno kategorizira 90 % primerov. Stopnja uspešnosti vzorcev nam daje pomemben vpogled v kompleksnost preučevanih kognitivnih procesov in z njim povezanih mehanizmov kodiranja in procesiranja informacij v možganih. Uspešni vzorci pa obetajo uporabnost na različnih področjih, od “branja možganov” do podpore diagnostiki možganskih motenj in bolezni ter spremljanja učinkovitosti njihovega zdravljenja.
Seveda MVPA še zdaleč ni rešitev vseh težav in ne daje vseh odgovorov na izzive analize fMR podatkov. Tehnika s seboj prinaša svoje lastne izzive in težave ter je uporabna le za naslavljanje specifičnih vprašanj, zagotovo pa kaže, da je vrhunsko raziskovanje s pomočjo fMR daleč od rdečih peg v sliki možganov mrtvega lososa.
-
___
-
Bennett, C.M., Baird, A.A., Miller, M.B., Wolford, G.L. (2009). Neural correlates of interspecies perspective talking in the post-mortem Atlantic Salmon: An argument for multiple comparison correction. 15th Annual Meeting of the Organization for Human Brain Mapping, San Francisco, CA, June 18-23, 2009. ↩
-
Repovš, G. (2009). Slikanje možganov. V M. Polič (ured.) Podobe psihologije, str. 127-146. Ljubljana: Znanstvena založba Filozofske fakultete. ↩
-
Friston, K.J., Holmes, A., Poline, J-B., Price, C.J., Frith, C.D. (1996), Detecting activations in PET and fMRI: Levels of inference and power. Neuroimage, 40, 223-235. ↩
-
Genovese, R. C, Lazar, N. A. in Nichols, T. (2002). Thresholding of statistical maps in functional neuroimaging using the false discovery rate. Neuroimage, 15 (4), 870–878. ↩
-
Nichols, T. E. in Holmes, A. P. (2001). Nonparametric permutation tests for functional neuroimaging: a primer with ex- amples. Human Brain Mapping, 15 (1), 1–25. ↩
prof. dr. Grega Repovš
Oddelek za psihologijo
Filozofska fakulteta
Univerza v Ljubljani